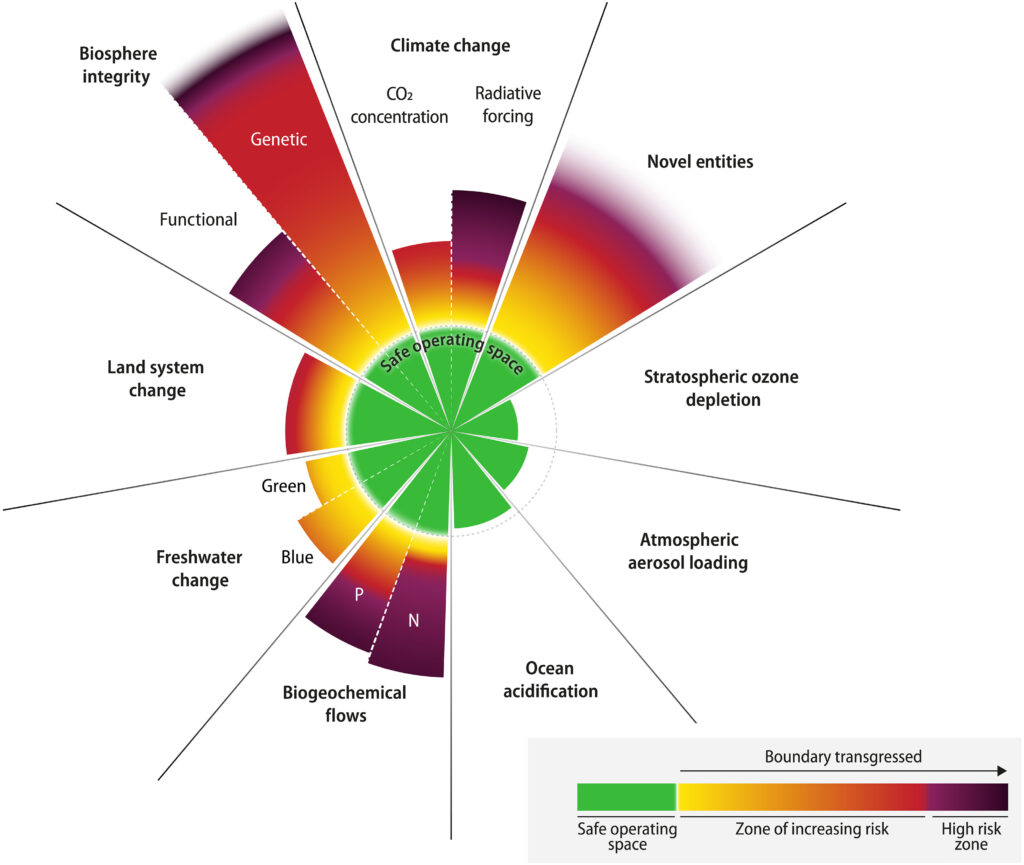
Insomma, secondo i ricercatori l’essere umano sta incidendo talmente tanto sul clima del Pianeta e sugli ecosistemi che abbiamo portato la Terra al di fuori dello “spazio sicuro operativo per l’umanità”.
In altre parole, in sei casi su nove le condizioni che negli ultimi 10mila anni hanno permesso all’essere umano di prosperare non sono più le stesse.
I livelli di ozono stratosferico si sono leggermente ripresi.
Il livello di trasgressione è aumentato per tutti i limiti precedentemente identificati come superati.
Poiché la produzione primaria guida le funzioni della biosfera del sistema Terra, l’appropriazione umana della produzione primaria netta viene proposta come variabile di controllo per l’integrità funzionale della biosfera. Anche questo confine viene oltrepassato.
Il quadro dei confini planetari si basa sulla scienza del sistema Terra . Identifica nove processi critici per il mantenimento della stabilità e della resilienza del sistema Terra nel suo complesso. Tutti sono attualmente pesantemente disturbati dalle attività umane. Il quadro mira a delineare e quantificare i livelli di perturbazione antropica che, se rispettati, permetterebbero alla Terra di rimanere in uno stato interglaciale “simile all’Olocene”. In tale stato, le funzioni ambientali globali e i sistemi di supporto vitale rimangono simili a quelli sperimentati negli ultimi 10.000 anni, anziché trasformarsi in uno stato senza analoghi nella storia umana. Questo periodo dell’Olocene, iniziato con la fine dell’ultima era glaciale e durante il quale si sono sviluppate l’agricoltura e le civiltà moderne, è stato caratterizzato da condizioni planetarie relativamente stabili e calde. Le attività umane hanno ora portato la Terra fuori dalla finestra di variabilità ambientale dell’Olocene.
Attualmente, le perturbazioni antropiche dell’ambiente globale vengono affrontate principalmente come se fossero questioni separate, ad esempio il cambiamento climatico, la perdita di biodiversità o l’inquinamento. Questo approccio, tuttavia, ignora le interazioni non lineari di queste perturbazioni e i conseguenti effetti aggregati sullo stato complessivo del sistema Terra. I confini planetari portano una comprensione scientifica degli impatti ambientali globali di origine antropica in un quadro che richiede di considerare lo stato del sistema Terra nel suo insieme.
Per più di 3 miliardi di anni, le interazioni tra la geosfera (flusso di energia e materiali non viventi nella Terra e nell’atmosfera) e la biosfera (tutti gli organismi/ecosistemi viventi) hanno controllato le condizioni ambientali globali. Lo stato del sistema Terra è cambiato in risposta alle forzanti generate da perturbazioni esterne (ad esempio, input di energia solare e attacchi di bolidi) o processi interni nella geosfera (ad esempio, tettonica a placche e vulcanismo) o biosfera (ad esempio, evoluzione della fotosintesi e crescita delle piante vascolari) . Queste forzanti sono state elaborate attraverso interazioni e feedback tra processi e sistemi all’interno del sistema Terra, modellandone la risposta complessiva, spesso complessa. Oggi, le attività umane con effetti su scala planetaria agiscono come ulteriore forzante sul sistema Terra. Così capace di alterare lo stato del sistema Terra. La struttura dei confini planetari formula limiti all’impatto dell’antroposfera sul sistema Terra identificando uno spazio operativo sicuro per l’umanità, basato su basi scientifiche, in grado di salvaguardare sia lo stato interglaciale della Terra che la sua resilienza.
Lo stato della Terra nell’Olocene è il riferimento di riferimento in questo contesto, poiché molti dei componenti che compongono la struttura dei confini planetari erano piuttosto stabili durante questo periodo. Questo è anche l’unico stato del sistema Terra che le civiltà hanno storicamente conosciuto. Il clima è una manifestazione di forzanti esterne, ad esempio l’attività solare, i cicli orbitali e le interazioni tra i componenti del sistema terrestre e la temperatura superficiale media globale varia di soli ± 0,5 ° C (9 (2 σ) (vedere i Materiali supplementari Neolitico [~9000 prima del presente (BP) ] fino alla Rivoluzione Industriale. Anche i biomi sulla Terra sono rimasti in gran parte stabili negli ultimi 10.000 anni, con una produzione primaria netta terrestre globale (NPP) preindustriale che variava di non > 55,9 ± 1,1 miliardi di tonnellate (Gt) di C anno −1 . dati confermano che anche i livelli di precipitazione globale preindustriale erano stabili, in particolare dalla metà dell’Olocene in poi. Questi dati forniscono un forte supporto per l’utilizzo dell’Olocene come stato di riferimento dei confini planetari per un pianeta stabile e resiliente.
Tutti i confini individuali del quadro adottano quindi le condizioni preindustriali dell’Olocene come riferimento per valutare l’entità delle deviazioni antropogeniche. I dati disponibili e lo stato delle conoscenze derivanti dall’analisi e dalla modellazione dei componenti del quadro dettano i metodi per la derivazione e la quantificazione dei singoli confini e dei relativi guardrail precauzionali. Nonostante i vincoli sui dati, sono stati compiuti sforzi per identificare variabili di controllo adeguate per tutti i confini, insieme a prove di quanto la perturbazione porti alla generazione di impatti o interazioni/feedback alterati che possono potenzialmente causare cambiamenti irreversibili ai sistemi di supporto vitale della Terra. L’attenzione è sempre rivolta al sistema Terra piuttosto che alla scala regionale, anche quando le prove utilizzate per stabilire i confini provengono da studi regionali. In questi casi,
Il quadro dei confini planetari ha attirato una notevole attenzione scientifica e sociale, ispirando strategie e politiche di governance a tutti i livelli. Il quadro si evolve attraverso aggiornamenti apportati alla luce delle recenti conoscenze scientifiche. Qui riuniamo i progressi di diversi campi della scienza per aggiornare il quadro e lo stato dei suoi confini. Per la prima volta vengono proposti dei confini per tutte le singole componenti del quadro. Gli aggiornamenti dell’integrità funzionale della biosfera e dei limiti di carico dell’aerosol si basano sulle analisi qui presentate. Analisi recenti costituiscono la base per gli aggiornamenti del cambiamento delle acque dolci e dei nuovi confini delle entità. Scorso,
Componenti della struttura
Comprendere come i processi della biosfera, dell’antroposfera e della geosfera interagiscono tra loro è un prerequisito per sviluppare proiezioni affidabili delle possibili traiettorie future del sistema Terra. Tuttavia, una comprensione pienamente basata sui processi delle interazioni tra questi ambiti è ancora disponibile solo parzialmente. Il quadro dei confini planetari richiede una modellizzazione più profondamente integrata del sistema Terra riunendo le prove attualmente disponibili per i processi rilevanti e le loro interazioni provenienti da diverse discipline e fonti.
I nove confini rappresentano tutti componenti del sistema Terra criticamente influenzati dalle attività antropiche e rilevanti per lo stato generale della Terra. Per ciascuno dei confini, vengono scelte variabili di controllo per catturare l’influenza antropica più importante a livello planetario del confine in questione. Ad esempio, il cambiamento del sistema terrestre deriva da una miriade di attività umane, che alla fine si aggregano all’alterazione dei biomi. Da una prospettiva planetaria, tuttavia, durante l’Olocene, le foreste erano il bioma terrestre con il più forte accoppiamento funzionale al sistema climatico . Pertanto, la riduzione globale della superficie forestale viene adottata come variabile di controllo che rappresenta tutti i cambiamenti del sistema terrestre. Allo stesso modo, la variabile di controllo qui introdotta per la componente funzionale del confine di integrità della biosfera, l’appropriazione umana della centrale nucleare (HANPP), si concentra sulla capacità della biosfera nel suo insieme di fornire feedback funzionali al sistema Terra. Le variabili di controllo dovrebbero idealmente prestarsi alla determinazione empirica ed essere calcolabili per l’uso nelle proiezioni del sistema Terra (ad esempio, simulazione basata sui processi dei futuri cambiamenti nella copertura forestale), ove possibile.
Le posizioni dei confini non delimitano né prevedono spostamenti di soglia singolari nello stato del sistema Terra. Sono posti a un livello in cui le prove disponibili suggeriscono che un’ulteriore perturbazione del processo individuale potrebbe potenzialmente portare a un cambiamento planetario sistemico alterando e rimodellando radicalmente le dinamiche e i modelli spaziotemporali delle interazioni geosfera-biosfera e i loro feedback.
La zona di rischio crescente (in cui il sistema Terra perde caratteristiche simili all’Olocene) viene ora utilizzata per valutare lo stato dei confini superati piuttosto che la “zona di incertezza” poiché la demarcazione di questa zona si basa su qualcosa di più di ciò a cui si fa solitamente riferimento. come incertezza scientifica. Un ampio corpus di ricerche recenti fornisce prove evidenti a sostegno della conclusione che i confini del cambiamento climatico e dell’integrità della biosfera si trovano in una zona caratterizzata da rischi in rapido aumento e sistemicamente collegati. Ciò rafforza la logica alla base dell’utilizzo del principio di precauzione per fissare i confini planetari all’estremità inferiore della zona di rischio crescente. Ad esempio, per il confine planetario del cambiamento climatico, manteniamo il limite di 350 parti per milione (ppm) di CO 2 con la zona di rischio crescente che va da 350 a 450 ppm prima di raggiungere il rischio elevato. Ciò corrisponde approssimativamente a un intervallo di aumento della temperatura superficiale media globale compreso tra 1° e 2°C (assumendo scenari tradizionali su livelli non-CO 2forzante). La precauzione colloca il confine planetario all’inizio del rischio crescente (350 ppm ≈ 1°C), cioè leggermente al di sotto dell’obiettivo di 1,5°C identificato nell’Accordo di Parigi. L’obiettivo di 1,5°C è un obiettivo che la scienza dimostra sempre più essere associato al rischio sostanziale di innescare grandi cambiamenti irreversibili e che il superamento di punti critici non può essere escluso anche con aumenti di temperatura più bassi . In riconoscimento della resilienza tampone del sistema Terra, la maggior parte dei confini sono tuttavia fissati a valori superiori al loro intervallo osservato attraverso l’Olocene fino alla Rivoluzione Industriale (per CO 2 ≈ 280 ppm) (vedere i Materiali Supplementari). La stabilità e la gamma caratteristica di variabilità degli stati del sistema terrestre interglaciale nel paleoclima del Pleistocene e la modellazione del sistema Terra suggeriscono che il sistema Terra probabilmente rimarrebbe in uno stato stabile, simile all’Olocene, se tutti i confini fossero rispettati nonostante siano almeno temporaneamente fuori dall’involucro della variabilità dell’Olocene.
La distinzione tra zone di rischio “crescente” e “alto” non può essere definita con precisione. Si stanno accumulando prove che l’attuale livello di trasgressione dei confini ha già portato il sistema Terra oltre una zona “sicura”. Tuttavia, manca ancora una teoria completa e integrata, supportata da osservazioni e studi di modellizzazione, in grado di identificare quando potrebbe verificarsi una transizione da un livello di rischio crescente a uno con rischi molto elevati e pericolosi di perdere uno stato del sistema Terra simile all’Olocene. Pertanto, in questo caso viene adottato l’approccio “tizzone ardente” introdotto dal Gruppo intergovernativo sui cambiamenti climatici (IPCC) per rappresentare le transizioni graduali dai rischi moderati (gialli) a quelli elevati (rosso) a molto elevati (viola).
RISULTATI
Integrità della biosfera
Una miriade di interazioni con la geosfera rendono la biosfera una componente costituzionale del sistema Terra e un fattore importante nella regolazione del suo stato. Il funzionamento planetario della biosfera si basa in ultima analisi sulla sua diversità genetica, ereditata dalla selezione naturale non solo durante la sua storia dinamica di coevoluzione con la geosfera, ma anche sul suo ruolo funzionale nella regolazione dello stato del sistema Terra. La diversità genetica e la funzione planetaria, ciascuna misurata attraverso adeguati proxy, sono quindi le due dimensioni che costituiscono la base di un confine planetario per l’integrità della biosfera. Come applicato qui, “integrità” non implica un’assenza di cambiamento della biosfera ma, piuttosto, un cambiamento che preservi il carattere dinamico e adattivo complessivo della biosfera.
Rockstrom et al. ( 1 ) hanno definito il limite planetario per il cambiamento della diversità genetica come il massimo tasso di estinzione compatibile con la preservazione della base genetica della complessità ecologica della biosfera. Manteniamo il livello limite di <10 E/MSY (estinzioni per milione di specie-anno). La variabile di controllo del tasso di estinzione è difficile da applicare in contesti operativi, ma stanno emergendo dati e metodi per valutare direttamente la componente di diversità genetica dell’integrità della biosfera [( 23) e i materiali supplementari]. Sebbene il tasso di estinzione di base (e dell’evoluzione di nuove specie) sia altamente variabile e difficile da quantificare con sicurezza nel corso del tempo geologico, si stima che l’attuale tasso di estinzione delle specie sia almeno da decine a centinaia di volte superiore al tasso medio negli ultimi 10 milioni di anni e sta accelerando ( 24 ). Fissiamo conservativamente il valore attuale per il tasso di estinzione a >100 E/MSY ( 24 – 26 ). Delle circa 8 milioni di specie animali e vegetali, circa 1 milione sono a rischio di estinzione ( 16 ) e oltre il 10% della diversità genetica di piante e animali potrebbe essere andato perduto negli ultimi 150 anni ( 23). Pertanto, la componente genetica del confine di integrità della biosfera viene notevolmente superata ( Fig. 1 e Tabella 1 ).
In precedenza, Steffen et al. ( 2 ) ha proposto di utilizzare l’indice di integrità della biodiversità (BII) ( 27 ), un sistema di misurazione basato empiricamente sugli impatti umani sull’abbondanza della popolazione, come indicatore provvisorio dell’integrità funzionale della biosfera. È stato notato, tuttavia, che il collegamento di BII alle funzioni del sistema Terra rimane poco compreso e BII non può essere direttamente collegato ai flussi biogeochimici ed energetici planetari rilevanti per stabilire lo stato del sistema Terra. Inoltre, BII si basa sulla raccolta di dati da parte di esperti per stimare i cambiamenti temporali nell’abbondanza/distribuzione delle specie, e questa conoscenza non è immediatamente disponibile per molte regioni, compresi gli oceani. Martin et al. ( 28) hanno recentemente suggerito che il BII riflette solo parzialmente gli impatti umani sul sistema Terra.
Pertanto ora sostituiamo questa metrica con un proxy calcolabile per l’energia fotosintetica e il flusso di materiali nella biosfera ( 29 ), cioè la produzione primaria netta (NPP), e definiamo la componente funzionale del confine di integrità della biosfera come limite all’appropriazione umana di la centrale nucleare della biosfera (HANPP) come frazione della centrale nucleare dell’Olocene. La centrale nucleare è fondamentale sia per gli ecosistemi che per le società umane poiché ne supporta il mantenimento, la riproduzione, la differenziazione, il networking e la crescita. I biomi dipendono dal flusso di energia associato alla centrale nucleare per mantenere le loro funzioni ecologiche planetarie come parti integranti del sistema Terra. I flussi di energia basati sulle centrali nucleari nelle società umane non dovrebbero quindi compromettere sostanzialmente il flusso di energia verso la biosfera ( 30). Il proxy integra le dimensioni dell’integrità della biosfera basate sulla diversità, coperte dalla componente genetica, che coglie l’importanza della variabilità negli organismi viventi per il funzionamento degli ecosistemi. L’idoneità di NPP e HANPP per definire un confine planetario è stata precedentemente discussa da Running ( 31 ) e Haberl et al. ( 32 ).
Determiniamo che la centrale nucleare dell’Olocene della biosfera terrestre è stata di 55,9 Gt di anno C −1 (2σ) ed estremamente stabile, variando di non più di ±1,1 Gt di anno C −1 nonostante le variazioni regionali nel tempo (vedere i Materiali Supplementari). Le nostre analisi dei modelli suggeriscono che la centrale nucleare aveva ancora un livello simile all’Olocene nel 1700 (56,2 Gt di anno C −1 per la potenziale vegetazione naturale e 54,7 Gt di anno C −1 quando si tiene conto dell’uso del suolo). Entro il 2020, il potenziale naturale della centrale nucleare sarebbe salito a 71,4 Gt di C anno −1 a causa della fertilizzazione con carbonio, una risposta di disequilibrio della fisiologia delle piante terrestri all’aumento antropogenico della CO 2concentrazione nell’atmosfera, mentre la centrale nucleare effettiva era di 65,8 Gt di C anno −1 a causa degli effetti di riduzione della centrale nucleare dell’uso globale del suolo (vedere i materiali supplementari).
HANPP designa sia la raccolta che l’eliminazione o l’alterazione (per lo più riduzione) del potenziale naturale NPP ( 32 ), principalmente attraverso l’agricoltura, la selvicoltura e il pascolo. L’HANPP terrestre può essere stimato sia come frazione della potenziale centrale nucleare naturale [15,7% nel 1950 e 23,5% nel 2020; dedotto da ( 33 ) e dai Materiali Supplementari] e della NPP media dell’Olocene (30% o 16,8 Gt di C anno −1nel 2020; vedere i materiali supplementari). Sosteniamo che un confine planetario basato sulla NPP che limiti l’HANPP dovrebbe essere fissato in relazione alla NPP media dell’Olocene preindustriale e non all’attuale potenziale NPP naturale. Questo perché l’aumento globale della centrale nucleare dovuto alla fertilizzazione del carbonio di origine antropica costituisce una risposta di resilienza del sistema Terra che smorza l’entità del riscaldamento di origine antropica. Pertanto, il contributo della centrale nucleare ad un deposito di carbonio associato alla fertilizzazione con CO 2 dovrebbe essere protetto e sostenuto piuttosto che considerato disponibile per la raccolta. In alcune regioni amazzoniche si osservano già, ad esempio, in alcune regioni dell’Amazzonia ( 34) e le foreste dell’Europa settentrionale.
Poiché l’NPP è la base del flusso di energia e materiali che è alla base del funzionamento della biosfera ( 30 ), sosteniamo che l’attuale impatto su scala planetaria dell’HANPP si riflette nell’osservazione che i principali indicatori dello stato della biosfera mostrano ampi e preoccupanti diminuzioni ultimi decenni ( 16). Ciò suggerisce che l’attuale HANPP è ben oltre un limite planetario precauzionale volto a salvaguardare l’integrità funzionale della biosfera e probabilmente si trova già nella zona ad alto rischio. Pertanto, abbiamo provvisoriamente fissato la componente funzionale del confine planetario dell’integrità della biosfera all’appropriazione umana del 10% della centrale nucleare media dell’Olocene preindustriale, spostandoci nella zona ad alto rischio al 20%. Il confine così definito fu oltrepassato alla fine del XIX secolo, un periodo di notevole accelerazione nell’uso del territorio a livello globale ( 35 ) con forti impatti sulle specie ( 27 ), che già fece sorgere le prime preoccupazioni sugli effetti di questa trasformazione del territorio su larga scala.
Pertanto, mentre il problema del riscaldamento climatico è diventato evidente negli anni ’80, i problemi legati all’integrità funzionale della biosfera dovuti all’uso del territorio da parte dell’uomo sono iniziati un secolo prima. A partire dagli anni ’60, la crescita della popolazione globale e dei consumi ha ulteriormente accelerato l’uso del territorio, spingendo il sistema ulteriormente nella zona di crescente rischio. L’HANPP ha sempre sostenuto il bisogno dell’umanità di cibo, fibre e foraggio, e continuerà ad essere così anche in futuro, così come per le società sostenibili. La centrale nucleare necessaria per sostenere le società future deve, tuttavia, essere generata sempre più attraverso una produzione aggiuntiva di centrale nucleare al di sopra della linea di base dell’Olocene, escludendo la centrale nucleare generata per i pozzi di assorbimento del carbonio su base biologica. Nutrire 10 miliardi di persone, ad esempio,36 ).
Per sviluppare una base più profonda per il confine planetario basato su HANPP per l’integrità funzionale della biosfera, abbiamo bisogno di una migliore comprensione di come le dinamiche ecologiche generano le funzioni della biosfera nel sistema Terra. L’analisi della centrale nucleare dovrebbe essere spazialmente esplicita e arricchita da parametri computabili di destabilizzazione ecologica dovuta alle pressioni sul clima e sull’uso del territorio, ad esempio, un parametro di disgregazione biogeochimica ( 37 ).
L’HANPP può essere quantificato anche per i sistemi marini. Circa due terzi dell’area oceanica in cui l’HANPP è >10% si trovano al di sopra delle aree poco profonde della piattaforma ( 38 ) dove gli ecosistemi sono più intensamente sfruttati. A livello regionale, le catture di pesce superano le soglie di sfruttamento sostenibile ( 39). Tuttavia, a differenza della terra, dove la maggior parte dell’HANPP si presenta sotto forma di materiale vegetale, cioè al livello trofico più basso, l’HANPP nell’oceano tende ad avvenire a livelli trofici più elevati. Ciò significa che mentre l’HANPP riduce la quantità assoluta di energia disponibile a livelli trofici più elevati sulla terra, gran parte dell’energia fissata attraverso l’NPP viene utilizzata negli ecosistemi marini prima che si verifichi l’HANPP. Quando l’abbondanza di organismi ai livelli trofici più alti viene ridotta, i cambiamenti nella struttura dell’ecosistema marino possono modificare il flusso di energia in questi ecosistemi ( 40). Pertanto, nel regno marino, l’HANPP probabilmente modifica i flussi piuttosto che la quantità di energia disponibile. Sono necessarie maggiori informazioni sugli impatti dell’HANPP nel regno marino per integrare la considerazione dei sistemi marini nel confine planetario dell’integrità funzionale della biosfera.
Cambiamento climatico
Le variabili di controllo del cambiamento climatico e i livelli limite vengono mantenuti ( 1 , 2 ). I fattori più importanti degli impatti antropici sul bilancio energetico della Terra sono l’emissione di gas serra e aerosol e i cambiamenti dell’albedo superficiale ( 17 ). Le variabili di controllo nel quadro sono le medie annuali della concentrazione atmosferica di CO 2 e la variazione della forzatura radiativa. Il limite planetario per la concentrazione atmosferica di CO 2 è fissato a 350 ppm e per la forzante radiativa a 1 W m −2 . Attualmente, la forzatura radiativa effettiva di origine antropica totale stimata è di 2,91 W m −2 [stima del 2022, rispetto a 1750 ( 17)], e la concentrazione atmosferica di CO 2 è pari a 417 ppm [valore medio annuo della superficie marina per il 2022 ( 41 )], ovvero più al di fuori dello spazio operativo sicuro su entrambe le misurazioni rispetto all’ultimo aggiornamento ( 2 ). Il limite di 350 ppm porterebbe a un livello inferiore di riscaldamento globale di origine antropica rispetto all’obiettivo di 1,5°C concordato a livello internazionale nell’Accordo di Parigi sul clima delle Nazioni Unite, ma è coerente con studi recenti (17, 18, 42) che suggeriscono la possibilità di un sistema Terra estremo . impatti anche con un riscaldamento di 1,5 °C , con rischi in aumento già nettamente al di sopra di 1°C di riscaldamento.
Entità nuove
La definizione di questo confine è ora limitata alle introduzioni antropiche veramente nuove nel sistema Terra. Questi includono sostanze chimiche e sostanze sintetiche (ad esempio, microplastiche, interferenti endocrini e inquinanti organici); materiali radioattivi mobilitati dall’uomo, compresi i rifiuti nucleari e le armi nucleari; e la modificazione umana dell’evoluzione, gli organismi geneticamente modificati e altri interventi umani diretti nei processi evolutivi. Le nuove entità fungono da indicatori geologici dell’Antropocene ( 5). Tuttavia, il loro impatto sul sistema Terra nel suo insieme rimane in gran parte non studiato. La struttura dei confini planetari riguarda solo la stabilità e la resilienza del sistema Terra, cioè non la salute umana o dell’ecosistema. Pertanto, rimane una sfida scientifica valutare quanto carico di nuove entità il sistema Terra tollera prima di spostarsi irreversibilmente in uno stato potenzialmente meno abitabile.
Centinaia di migliaia di sostanze chimiche sintetiche vengono ora prodotte e rilasciate nell’ambiente. Per molte sostanze, gli effetti potenzialmente ampi e persistenti derivanti dalla loro introduzione sui processi del sistema Terra, in particolare sull’integrità funzionale della biosfera, non sono ben noti e il loro utilizzo non è ben regolamentato. L’umanità è stata ripetutamente sorpresa dalle conseguenze indesiderate di questo rilascio, ad esempio, rispetto al rilascio di insetticidi come il DDT e all’effetto dei clorofluorocarburi (CFC) sullo strato di ozono. Per questa classe di nuove entità, quindi, l’unico spazio operativo veramente sicuro in grado di garantire il mantenimento di condizioni simili all’Olocene è quello in cui queste entità sono assenti a meno che i loro potenziali impatti rispetto al sistema Terra non siano stati attentamente valutati. Ciò implicherebbe che il limite planetario quantificato dovrebbe essere fissato a zero rilascio di composti chimici sintetici nell’ambiente aperto a meno che non siano stati certificati come innocui e siano monitorati. Questo è l’obiettivo fissato dal Protocollo di Montreal per quanto riguarda le sostanze che si sono rivelate dannose perché contribuiscono alla riduzione dello strato di ozono.
Nella loro analisi di varie strategie per stabilire un confine planetario per nuove entità, Persson et al. ( 43) ha individuato come possibile variabile di controllo la quota di sostanze chimiche rilasciate soggette a valutazione e monitoraggio della sicurezza adeguati. Qui adottiamo questa metrica. Il limite planetario viene quindi fissato al rilascio nel sistema terrestre dello 0% di materiali sintetici non testati. Quando i materiali sintetici rilasciati nell’ambiente vengono testati a fondo, il conseguente rischio di effetti dannosi viene ridotto. Certo, questo approccio presenta dei punti deboli: la disponibilità dei dati è incompleta; gli studi sulla sicurezza spesso si concentrano su una tossicità ben definita e non catturano gli “effetti cocktail” delle miscele chimiche nell’ambiente né i loro effetti in condizioni specifiche. La percentuale di sintetici non testati rilasciati a livello globale è sconosciuta. Tuttavia, Persson et al. ( 43) riferiscono che per le sostanze chimiche attualmente registrate ai sensi del regolamento UE sulla registrazione, valutazione, autorizzazione e restrizione delle sostanze chimiche (REACH) (un piccolo sottoinsieme dell’universo chimico), circa l’80% di queste sostanze chimiche è in uso da almeno 10 anni senza ancora sottoposto a una valutazione di sicurezza. Allo stesso modo, pochi studi sulla sicurezza considerano i potenziali effetti sul sistema Terra. Con una percentuale così enorme di sostanze chimiche non testate rilasciate nell’ambiente, un nuovo confine di entità definito in questo modo viene chiaramente violato. Persson et al. ( 43 ) non ha individuato né quantificato un limite planetario unico per le nuove entità, ma ha comunque concluso che lo spazio operativo sicuro è attualmente oltrepassato.
Distruzione dell’ozono stratosferico
La riduzione dell’ozono stratosferico è un caso speciale legato al rilascio antropogenico di nuove entità in cui i composti gassosi di alocarburi provenienti dall’industria e da altre attività umane rilasciati nell’atmosfera portano a una riduzione duratura dello strato di ozono terrestre. Il limite per lo spazio operativo sicuro è fissato a 276 unità Dobson (DU), consentendo cioè una riduzione <5% rispetto al livello preindustriale di 290 DU, valutato in base alla latitudine ( 1 ) . Dopo la ratifica del Protocollo di Montreal nel 1987, la tendenza e l’entità globale della riduzione dell’ozono hanno registrato una leggera ripresa ( 44 , 45). L’attuale stima globale (2020) è di 284 DU (vedere i Materiali Supplementari). Pertanto, il perturbamento umano della riduzione dell’ozono stratosferico è diminuito e ora rientra nello spazio operativo sicuro. Il limite per la riduzione dell’ozono viene attualmente oltrepassato solo nell’Antartide e alle alte latitudini meridionali e solo nei tre mesi della primavera australe ( 45 ).
Cambio d’acqua dolce
Per riflettere in modo completo le modifiche antropogeniche delle funzioni del sistema terrestre di acqua dolce, questo confine è rivisto per considerare i cambiamenti attraverso l’intero ciclo dell’acqua sulla terra ( 46 – 48 ). Qui utilizziamo il flusso del flusso come proxy per rappresentare l’acqua blu (superficiale e sotterranea) e l’umidità del suolo nella zona delle radici per rappresentare l’acqua verde (acqua disponibile per le piante) ( 46 – 48 ). Le variabili di controllo sono definite come la percentuale di superficie annuale globale libera dai ghiacci con deviazioni dell’umidità del suolo nella zona del flusso d’acqua/zona radicale rispetto alla variabilità preindustriale ( 46 , 48). La nuova componente acqua verde tiene conto direttamente della regolazione idrologica degli ecosistemi terrestri, del clima e dei processi biogeochimici ( 48 ), mentre la componente acqua blu tiene conto della regolazione dei fiumi e dell’integrità dell’ecosistema acquatico ( 46 ). Inoltre, questo confine ora cattura gli impatti sul sistema Terra sia degli aumenti che delle diminuzioni dell’acqua su scala mensile e include i loro modelli spaziali (vedere i Materiali Supplementari).
Le variabili di controllo descrivono le deviazioni rispetto allo stato preindustriale (qui, 1661–1860), determinate inizialmente alla scala delle celle della griglia di 30 arco-min e successivamente aggregate in un valore annuale globale. Sia per le variabili di controllo dell’acqua blu che per quelle verdi, i confini sono fissati al 95° percentile della variabilità preindustriale, cioè la variabilità della percentuale di superficie terrestre globale con deviazioni [~10% per l’acqua blu e ~11% per l’acqua verde; ( 46) e i materiali supplementari]. Partiamo dal presupposto che le condizioni preindustriali siano rappresentative delle condizioni dell’Olocene a lungo termine e che una notevole deviazione da questo stato metta a rischio le funzioni del sistema terrestre di acqua dolce. In attesa di una valutazione completa degli impatti dei diversi livelli di trasgressione dei confini delle acque blu e verdi (ad esempio, ridotta capacità di sequestro del carbonio, regolazione del clima e perdita di biodiversità; vedere i materiali supplementari), le impostazioni dei confini sono preliminari e altamente precauzionali. Attualmente, circa il 18% (acque blu) e circa il 16% (acque verdi) della superficie terrestre globale presenta deviazioni di acqua dolce umida o secca ( 46 ). Pertanto, in contrasto con le precedenti valutazioni sui confini planetari ( 1 , 2) laddove è stata presa in considerazione solo la rimozione dell’acqua blu, questo nuovo approccio indica una sostanziale trasgressione del limite di cambiamento dell’acqua dolce. Le trasgressioni dei confini dell’acqua blu e verde si sono verificate un secolo fa, rispettivamente nel 1905 e nel 1929 ( 46 ). Pertanto, con la nuova definizione delle variabili di controllo, l’acqua dolce sarebbe stata considerata superata già al momento delle precedenti valutazioni dei confini planetari. La precedente variabile di controllo su scala globale indicherebbe comunque che l’uso di acqua dolce rimane nella zona sicura, anche con fonti di dati più recenti rispetto a quelle utilizzate in ( 1 , 2 ). Stime recenti del consumo globale di acqua blu ammontano a ~1700 km 3 anni −1 ( 49), cioè molto al di sotto del limite precedente fissato a 4000 km 3 anni −1 .
Carico di aerosol atmosferico
Gli aerosol hanno molteplici effetti fisici, biogeochimici e biologici nel sistema Terra, motivando la loro inclusione come confine planetario (vedi Materiali Supplementari). Il carico di aerosol antropogenico è aumentato ( 50 ). I cambiamenti dal periodo preindustriale per gli aerosol naturali (ad esempio, polvere del deserto, fuliggine da incendi) sono difficili da valutare a causa delle differenze dei modelli nel segno delle tendenze ( 51 ) , ma le prove osservative suggeriscono un raddoppio globale della deposizione di polvere dal 1750 ( 52 ). Al momento, il Sahara è la più grande regione di origine delle polveri al mondo [ad esempio, ( 53)], ma prima nell’Olocene, era un paesaggio ricoperto di vegetazione con molti laghi e zone umide (da 14.500 a 5.000 anni fa). Si ritiene che i cambiamenti nelle piogge monsoniche, che comportano feedback tra vegetazione, polvere e clima, abbiano posto fine al “Sahara verde”, portando a grandi spostamenti di insediamenti umani in parti dell’Africa e dell’Asia ( 54 ) .
La quantificazione del confine planetario del carico di aerosol è ostacolata dalle loro molteplici fonti naturali e causate dall’uomo, dalle differenze nella composizione chimica, dalla stagionalità e dalla durata atmosferica e dalla conseguente grande eterogeneità spaziale e temporale nella distribuzione e negli impatti climatici ed ecologici degli aerosol. Tuttavia, la profondità ottica dell’aerosol (AOD) fornisce una variabile di controllo generica per il carico di aerosol. L’AOD è una misura integrata della riduzione complessiva della luce solare che raggiunge la superficie terrestre causata da tutto l’assorbimento e la diffusione nella colonna d’aria verticale. Sulla base delle prove degli impatti di grandi AOD sulle precipitazioni regionali nell’Asia meridionale, Steffen et al. ( 2) stabiliscono un limite planetario regionale provvisorio di AOD = 0,25 (da 0,25 a 0,5) sulla base del fatto che valori AOD più elevati nelle regioni monsoniche probabilmente portano a precipitazioni significativamente inferiori, influenzando in definitiva l’integrità della biosfera. L’AOD medio annuo nell’Asia meridionale è attualmente compreso tra 0,3 e 0,35 ( 55 , 56 ). Il valore attuale per la regione della Cina orientale è 0,4 ( 55 ). Pertanto, il carico di aerosol in queste regioni ha probabilmente superato il confine definito a livello regionale, ma con elevata incertezza. Mancano dati e valutazioni degli impatti degli aerosol sul clima e sugli ecosistemi per determinare se questo confine definito a livello regionale sia applicabile altrove. La media globale dell’AOD attualmente è 0,14 ( 57), con livelli molto più elevati in alcune regioni e con gradienti molto forti dalla terra all’oceano aperto ( 56 ).
Oltre agli effetti diretti dell’AOD sul clima regionale e sulle precipitazioni, le asimmetrie dell’AOD tra gli emisferi settentrionale e meridionale possono influenzare più sistemi monsonici, come visto per il monsone dell’Africa occidentale (58) e il monsone indiano ( 59 , 60 ) . La differenza interemisferica nell’AOD influenza le precipitazioni monsoniche regionali spostando la posizione della zona di convergenza intertropicale ( 61 ). Grandi asimmetrie nella temperatura degli emisferi settentrionale e meridionale derivano dalle differenze nelle emissioni di aerosol naturali e antropogeniche, nella copertura del suolo e in altri fattori climatici ( 58 , 59 , 62 , 63) .). La forzante radiativa asimmetrica derivante dagli effetti degli aerosol porta ad un relativo raffreddamento dell’emisfero settentrionale e ad uno spostamento verso sud delle precipitazioni tropicali ( 64 ). La differenza AOD interemisferica e il suo impatto sulle precipitazioni tropicali e sulla disponibilità di acqua sono sensibili alla dimensione delle particelle e alla distribuzione latitudinale e altitudinale degli aerosol ( 65 ). Studi sulle interazioni aerosol-clima a seguito di eruzioni vulcaniche ( 66) indicano che le precipitazioni monsoniche nell’emisfero settentrionale sono indebolite quando l’AOD dell’emisfero settentrionale è più elevato e la differenza dell’AOD interemisferico è maggiore e aumenta quando vengono emessi più aerosol nell’emisfero meridionale (differenza dell’AOD interemisferico minore). Questa comprensione è sostanzialmente coerente con la diminuzione delle precipitazioni medie tropicali dopo le grandi eruzioni vulcaniche nelle osservazioni e nei modelli climatici globali ( 67 ). L’IPCC AR6 ha valutato che le diminuzioni osservate nelle precipitazioni monsoniche terrestri globali dagli anni ’50 agli anni ’80 sono in parte attribuite alle emissioni di aerosol dell’emisfero settentrionale causate dall’uomo, quindi a una differenza interemisferica relativamente maggiore ( 17). Oltre agli aerosol vulcanici, le dinamiche dei monsoni e le precipitazioni regionali associate rispondono anche ai cambiamenti negli aerosol di origine antropica (vedere i Materiali Supplementari).
Proponiamo quindi la differenza interemisferica media annuale dell’AOD come variabile di controllo definita a livello globale per il carico di aerosol. L’attuale differenza interemisferica è di ~0,076 ± 0,006 (media ± SD), sulla base di 12 stime osservative, raggiungendo ~0,1 nella primavera e nelle estati boreali, a causa dell’aumento stagionale delle tempeste di polvere che dominano nell’emisfero settentrionale ( 55 ) . . Il valore medio annuo preindustriale è stimato pari a ~0,03, sulla base di analisi multimodello ( 68), indicando un aumento della differenza AOD interemisferica di ~ 0,04 nell’era industriale. L’attuale differenza AOD interemisferica è coerente con gli inventari delle emissioni del Coupled Model Intercomparison Project 6 (CMIP6) che mostrano più aerosol di origine antropica nell’emisfero settentrionale, con proiezioni future che suggeriscono una diminuzione dell’asimmetria (69 ) .
Assegniamo un valore limite planetario di 0,1 per la differenza interemisferica media annuale dell’AOD, con elevata incertezza sulla zona di rischio crescente, da 0,1 a 0,25. Nel fissare questo confine, notiamo che gli impatti del carico di aerosol sui sistemi monsonici tropicali sono già visibili oggi, e l’impatto non è limitato solo alle precipitazioni ma influenza anche il clima regionale in modo più ampio. L’interazione aerosol-nube potrebbe esacerbare gli effetti dell’asimmetria AOD. Il contributo delle interazioni aerosol-nube all’asimmetria emisferica della radiazione ad onde corte riflessa non è chiaro. Prendiamo ad esempio l’attuale intervallo di forzatura radiativa efficace dell’aerosol antropogenico per i giorni nostri che è stato segnalato essere compreso tra -1,6 e -0,6 W m -2nella media globale per l’intervallo di confidenza dal 16 all’84%, con le interazioni aerosol-nuvole come una delle principali fonti di incertezza ( 51 ). Sono già evidenti anche altri effetti su larga scala degli aerosol, come gli impatti sulla qualità dell’aria sugli ecosistemi terrestri e marini ( 17 , 70 ). Gli aerosol biogenici non sono stati considerati, nonostante il loro ruolo nei feedback nel sistema Terra. È necessaria una migliore comprensione sistemica e quantitativa degli effetti idroclimatici, ecologici e biogeochimici della forzatura asimmetrica dell’aerosol per affinare il limite di carico dell’aerosol.
Acidificazione degli oceani
La variabile di controllo utilizzata è la concentrazione di ioni carbonato nell’acqua di mare superficiale (in particolare, Ω arag , lo stato di saturazione medio dell’oceano superficiale globale rispetto all’aragonite). La quantificazione del confine originale [≥80% della media preindustriale globale di Ω arag di 3,44 ( 1 )] viene mantenuta. Una stima recente fissa l’attuale Ω arag a ~2,8 ( 71 ) (vedi Materiali Supplementari), circa l’81% del valore preindustriale. Pertanto, l’acidificazione degli oceani di origine antropica si trova attualmente ai margini dello spazio operativo sicuro e la tendenza sta peggiorando poiché le emissioni di CO2 di origine antropica continuano ad aumentare.
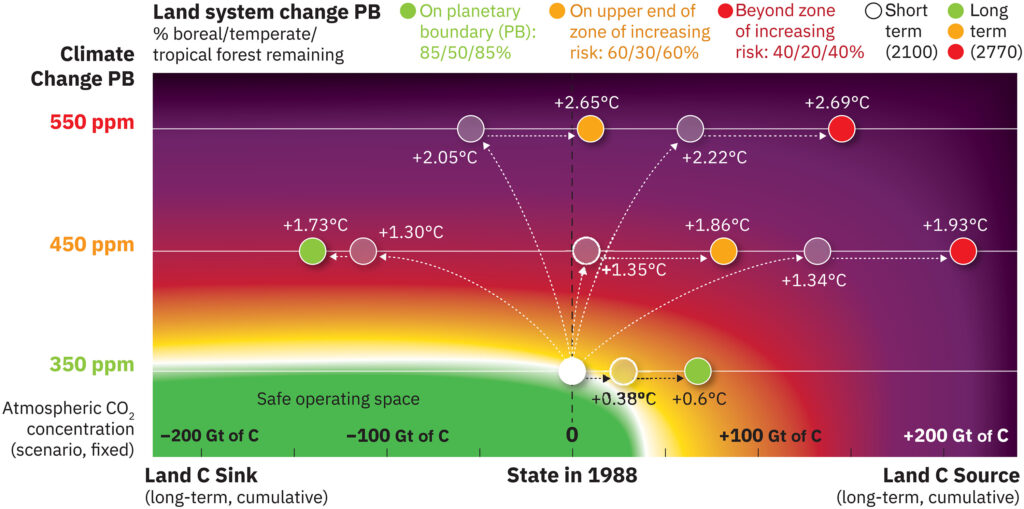
Cambiamento del sistema fondiario
Questo confine si concentra sui tre principali biomi forestali che a livello globale svolgono il ruolo più importante nel guidare i processi biogeofisici ( 2 ), vale a dire tropicale, temperato e boreale. La variabile di controllo rimane la stessa: la copertura forestale rimanente rispetto alla superficie forestale potenziale nell’Olocene ( 2 ). Le posizioni limite rimangono all’85%/50%/85% per le foreste boreali/temperate/tropicali (cfr. Tabella 1 e Materiali Supplementari). Sulla base delle mappe di classificazione della copertura del suolo del 2019 derivate da osservazioni satellitari ( 72), lo stato attuale dei biomi regionali è simile a quello del 2015 sebbene, per la maggior parte delle regioni, la quantità di deforestazione sia aumentata dal 2015 (vedi Materiali Supplementari). La conversione dell’uso del territorio e gli incendi stanno causando rapidi cambiamenti nell’area forestale ( 73 , 74 ), e la deforestazione della foresta tropicale amazzonica è aumentata a tal punto da aver oltrepassato il confine planetario ( Tabella 1 ). I cambiamenti nella metodologia e nella tecnologia utilizzata per stimare la copertura forestale dal 2015 potrebbero influenzare le differenze a livello di bioma qui riportate rispetto all’ultimo aggiornamento ( 2 ). Ciononostante, non vi sono dubbi sul fatto che la superficie forestale globale continui a diminuire ( 74 ).